Dehua Pei, PhD, Professor, Ohio State University
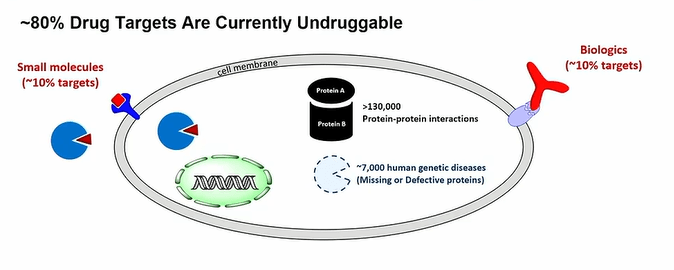
Dr. Dehua Pei introduced his talk by sharing that more than 80% of the genetic diseases are undruggable, and arguing we need new classes of drugs such as intracellular biologics that get into the intracellular space to perform their functions such as enzymes, antibodies, endonucleases, RNA, peptides, antisense oligonucleotides and so on. The main point of his talk is how these molecules get into the intracellular space and how that enables targeted delivery and therapeutic treatment.
Dr. Pei started by using the example of Enhanced Green Fluorescent Protein (EGFP). In the research Dr. Pei conducted, when the loop structure of EGFP that had no functional importance was replaced by a cell penetrating peptide motif R4W3, it enabled proteins to get into the cell. Replacement of the loop structure by this 7-aa motif caused an intracellular fluorescent signal. Pei’s team also tested the same strategy on another protein, PTP1B enzyme which hydrolyzes phosphotyrosine in cytosol and showed that engineered PTP1B reduced tyrosine
phosphorylation in a dose-dependent manner while WT PTP1B could not. He also mentioned some drawbacks to this approach. First, the addition of a new loop sequence might disrupt 3D structure, but it is possible to make predictions via tools such as AlphaFold [1] beforehand. Another difficulty is the requirement of trying different loop sequences and optimization of the protocols to find the molecule needed. He defined the required features of the protein as follows:
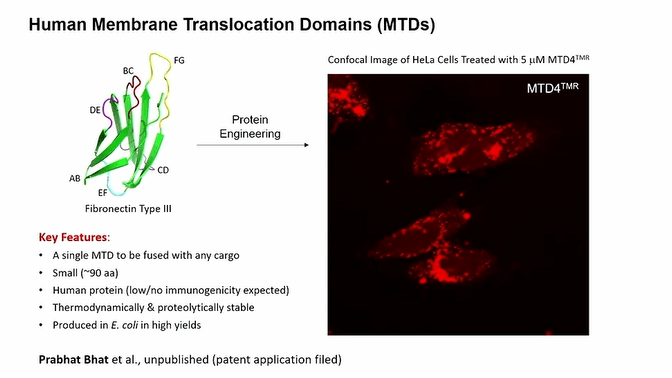
In the light of these points, Pei’s team focused on a Fibronectin III domain protein, membrane translocation domains (MTDs) which is small (approximately 90 aa in size), suitable to be fused with any cargo, stable and not immunogenic. He shared data indicating that when HeLa cells were treated with MTD labeled with a dye, the proteins entered into the cells and gave signal from intracellular space. He also explained possible designs based on MTD including therapeutic proteins, target domains for targeted delivery, E3 ligase and target domain for tissue specific protein degradation or just a drug molecule.
Dr. Pei underlined the importance of PROTAC strategy [2] in therapeutic development. PROTAC is a bifunctional molecule containing one molecule for binding to target and another molecule for recruiting E3 ligase so that the target is ubiquitinated and degraded by a proteasome. The major limitation of this strategy is the need for a small molecule that binds to the protein of interest (POI). He stated that if this is achieved by proteins, we will be able to target and degrade any target POI. The data they generated indicated that the chimeric protein consisting of E3 ligase, Ras Binding Domain (RBD) and MT4 caused dose-dependent degradation of Ras and Ras knockdown cells displayed a loss of viability.
As an alternative application, Dr. Pei talked about the potential of cell-permeable proteins for sustainable agriculture. He mentioned harpin proteins which are produced by bacteria stimulate plant growth, and enhance tolerance to stress. Harpin protein is commercially available but due to poor cell penetration efficiency, a high concentration level is required. Dr. Pei said that once we could deliver these proteins into plant cells via MTD, there might be a 10-100 fold increase in the efficiency of the delivery leading to a reduction in cost. Another finding shared was that tomatoes treated with MTD4-HrpZ after fungal infection displayed resistance against fungal growth.
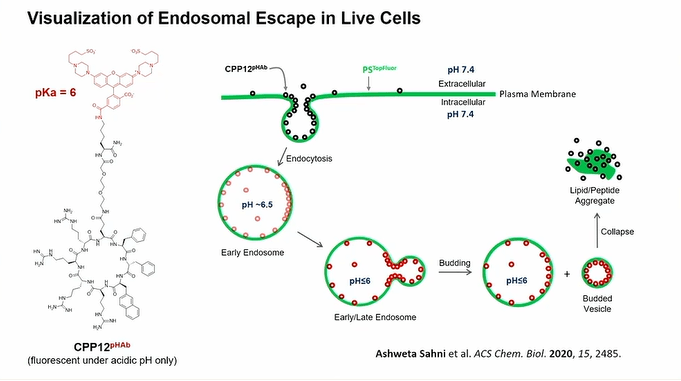
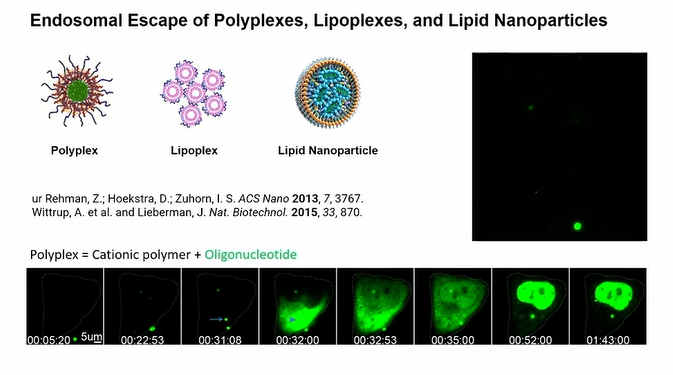
Following the applications, Dr. Pei explained how these biomolecules cross the cell membrane by focusing on cyclic cell-penetrating peptides (CCPP) consisting of four arginines and two hydrophobic residues as known as endosomal escape vehicles (EEVs). The mechanism they proposed involves CCPPs binding to a plasma membrane by probably interacting with phospholipids and then being taken into the early endosome through endocytosis. As the endosome matures, pH within the endosome becomes more acidic and CCPPs bind to the endosome membrane with higher affinity and start to cluster together to form CPP rich lipid domains which make budding structures. The budded vesicle is separated from the endosome, collapsed, and translocation of CPP molecules from extracellular space to cytosolic space take place. They explored this phenomenon using several endocytosis inhibitors and pH-sensitive dyes.
In the last part, Dr. Pei mentioned the structural features driving an effective VBC process and explained the high endosomal escape efficiency of CPP12 and CPM3 in comparison to Tat displaying poor endosomal escape and stated that CPP12 and CPM3 are amphipathic, have positively charged arginine and arginine-like moieties with hydrophobic groups and are structurally constrained by cyclization due to the benzene ring. He underlined the importance of having amphipathic structures of proper conformation as well as conformational rigidity that results in greater membrane-binding affinity.
References
[1] Database, A. F. P. S. (n.d.). Alphafold protein structure database. AlphaFold Protein Structure Database. Retrieved November 3, 2022, from https://alphafold.ebi.ac.uk/ [2] Qi, S.-M., Dong, J., Xu, Z.-Y., Cheng, X.-D., Zhang, W.-D., & Qin, J.-J. (1AD, January 1). PROTAC: An effective targeted protein degradation strategy for cancer therapy. Frontiers. Retrieved November 3, 2022, from https://www.frontiersin.org/articles/10.3389/fphar.2021.692574/full