A whitepaper by Genscript on exploring novel MOA and validating bi-specific antibodies against PD-1 and TIGIT
After the first wave of therapeutic antibody development towards immunology/ oncology, fewer and fewer new targets emerged in recent years.
Instead, people start to shift more attention onto novel modality and MOA against the same target. As a result, the bi-specific antibody has become one of the mainstream strategies.
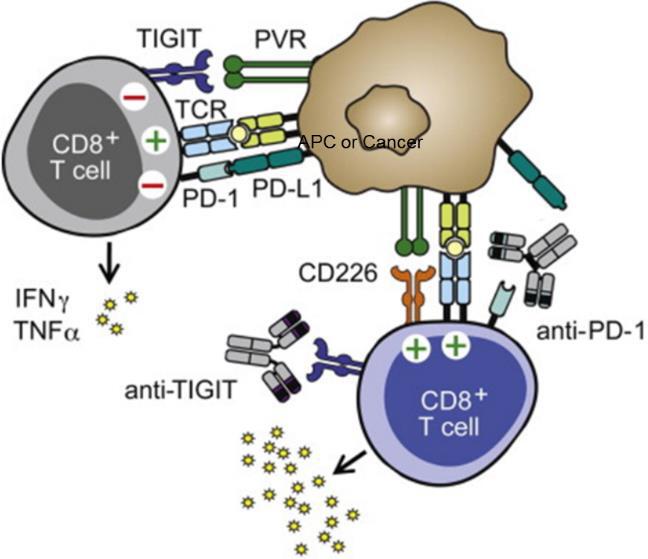
Both PD-1 and TIGIT are well-established drug targets for immunology/oncology therapy. TIGIT is expressed on the surface of activated T cells and NK cells as a marker of exhaustion, and its expression level is upregulated in a variety of cancers.
As an inhibitory checkpoint implicated in tumor immunosurveillance, it sends a suppressive signal through PVR (CD155) direct binding and competitive binding against CD226.
Similarly, PD-1 is also an immunosuppressive checkpoint.
Dual inhibition of these two targets may reverse T-cell exhaustion, improve cytotoxic T-cell effector function, and restore the potency of cytotoxic T cells and NK cells within a highly suppressive tumor microenvironment (figure 1).
Figure 1. Dual inhibition of cancel cell by TIGIT and PD-1 signaling pathway1
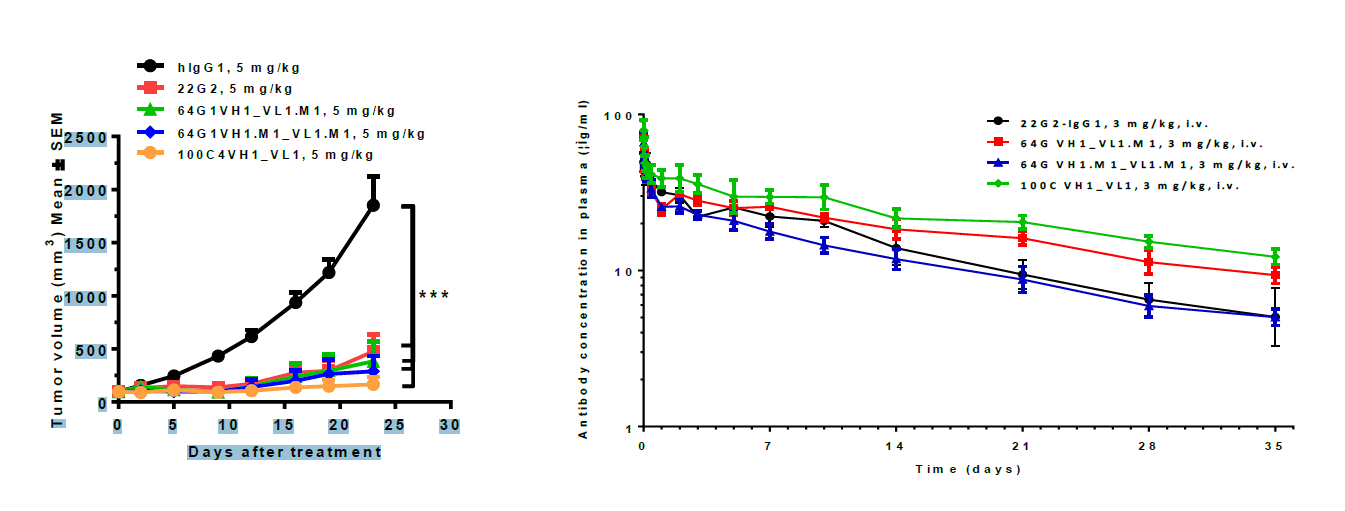
For TIGIT monotherapy, IgG is still considered the most popular format. By using different approaches, including hybridoma, single B cell cloning, and phage display, a panel of lead candidates was identified.
These antibodies showed similar or even better in vivo efficacy and PK profile compared to benchmark 22G2 (figure 2).
Figure 2. Comparison of in vivo efficacy and PK profile between lead candidates and benchmark (22G2)
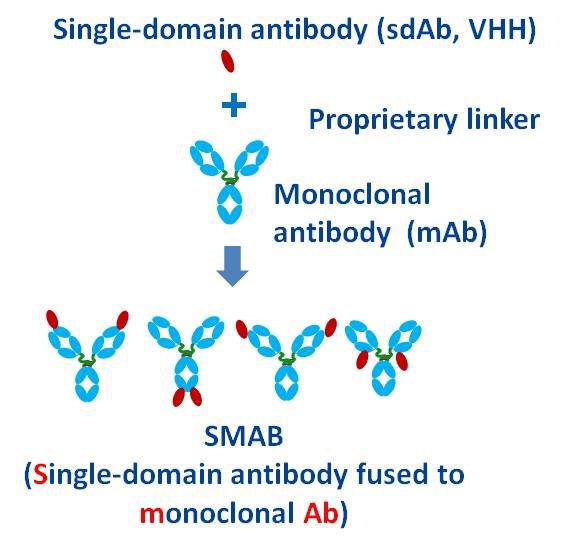
For TIGIT/PD-1 bi-specific antibody, we utilized the GenScript SMABody format (figure 3), which consists of VHH against TIGHT and IgG against PD-1.
The VHH antibodies were generated from Ilama immunization followed by phage display approach, while the anti-PD-1 IgG candidates were originated from immunized mice and hybridoma platform.
These two sets of antibody leads were evaluated by a single targeted assay, and one top candidate from each set was constructed into bi-specific SMABody BTP-11 and BTP-13.
In order to verify the retaining of activity after bi-specific construction, single target cellbased assays were conducted.
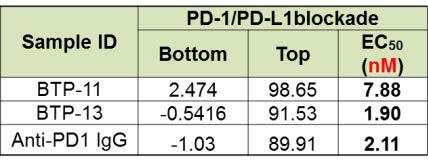
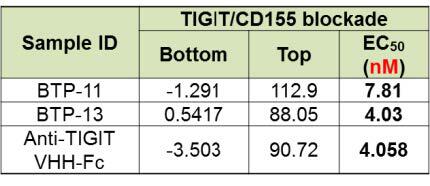
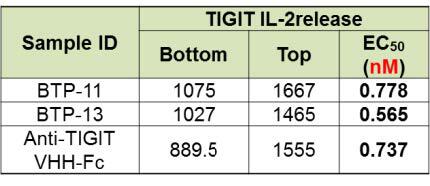
Both BTP-11 and BTP-13 showed similar efficacy as compared to its parental antibody in PD-1/PD-L1 blockade (figure 4, top), TIGIT/CD155 blockade (figure 4, middle), and TIGIT IL-2 release assay (figure 4, bottom).
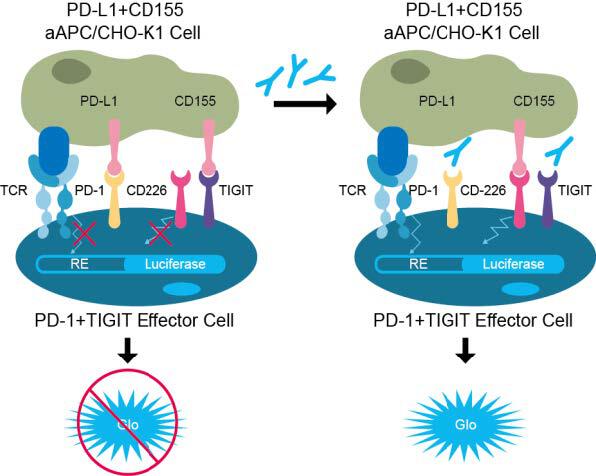
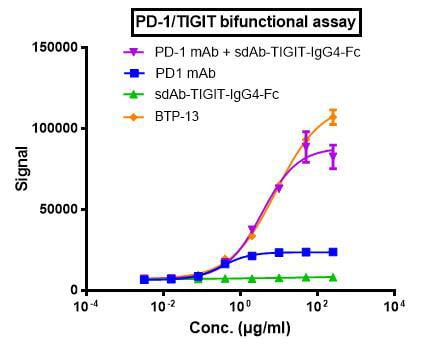
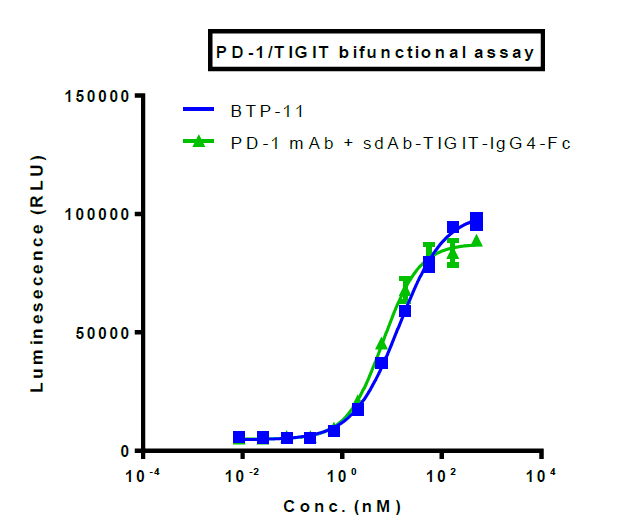
To further characterize these bi-specific antibodies under the scenario of both PD-1 and TIGIT, we built a dual-target reporter assay (figure 5, top), in which the simultaneous blocking of TIGIT and PD-1 signaling leads to the induction of luciferase.
In this assay, both BTP-11 and BTP-13 showed better efficacy compared to the combination of IgG and sdAb, and the single antibody groups demonstrated minimal activation of signaling (figure 5, bottom).
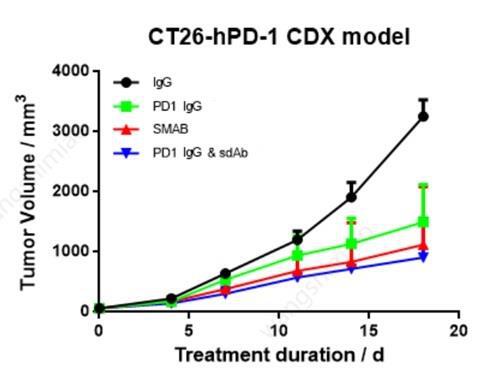
Furthermore, in vivo efficacy of top SMAB candidate was evaluated in mice CDX model, along with single target antibody and combination.
The bi-specific antibody demonstrated comparable inhibition as IgG+sdAb combo, and both of them showed much better response than negative control (IgG) and monoclonal antibody (PD-1 IgG).
Figure 3. Configuration of SMABody
Figure 4. Evaluation of bi-specific antibody by single target cell-based assay
Figure 5. In vitro efficacy evaluation by dual-target reporter assay
Figure 6. Comparison of in vivo efficacy between bi-specific SMAB vs. single target antibodies
In summary, several IgG candidates against TIGIT are already in the clinical stage. Exploring novel MOA and validating bi-specific antibodies against both PD-1 and TIGIT provides an alternative solution against established targets.
The GenScript SMABody bispecific format consists of IgG and sdAb by simple fusion. Its symmetrical structure eliminates the need of Fc/Fab engineering and significantly reduces the developability issues during the manufacturing stage.
[1] Johnston R , Comps-Agrar L , Hackney J , et al. The immunoreceptor TIGIT regulates antitumor and antiviral CD8(+) T cell effector function.[J]. Cancer Cell, 2014, 26(6):923-937.