Gyros Protein Technologies
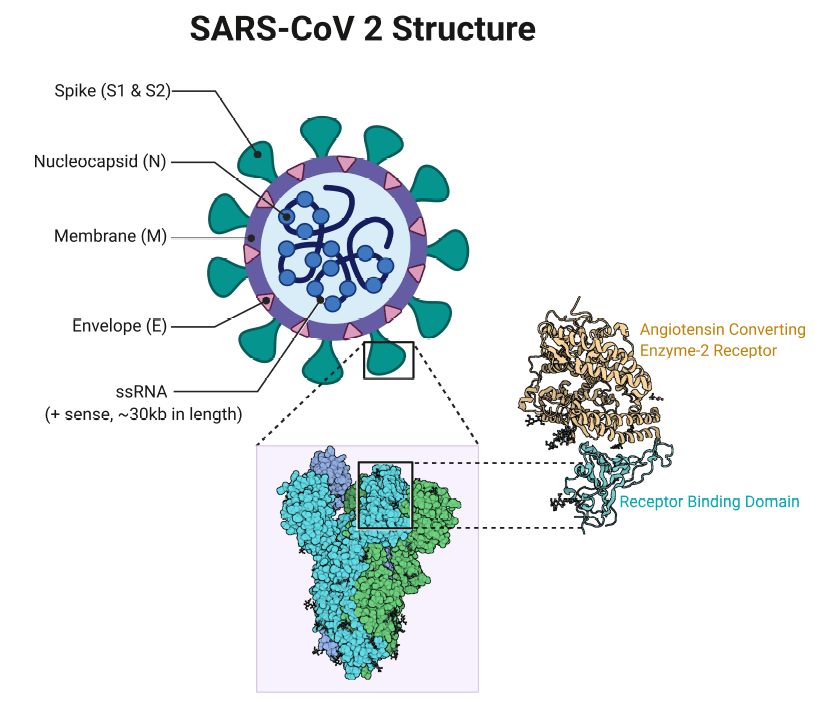
Figure 1. The structure of SARS-CoV-2. Contributed by Rohan Bir Singh, MD; made with Biorender.com. Included under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), from reference 10.
The COVID-19 pandemic caused by the novel coronavirus SARS-CoV-2 has triggered intense activity around the world and may well represent a paradigm shift in fighting infectious disease. For example, the SARS-CoV-2 genome was published on January 12, 2020, less than two weeks after the first cases were reported, and the first vaccine candidate entered human clinical testing just two months later – just one example of an enormous range of prototype therapeutics and vaccines being developed (1–3).
Peptide synthesis is one valuable component of the toolbox needed to quickly investigate the mechanism of infection, and peptides may also prove to have therapeutic properties or form the basis of a vaccine.
Therapeutic peptides have many advantages, including high activity, great chemical and biological diversity, and low toxicity. These and the relative ease and low cost of peptide manufacture compared to protein-based biologicals has meant that therapeutic peptides are now being used to treat a range of conditions, including metabolic diseases, cancer, cardiovascular, and infectious diseases.
The speed and flexibility of peptide synthesis is a major advantage when handling rapidly evolving conditions, such as neoantigen peptide-based vaccines for the individualized immunotherapy of certain forms of cancer. The dynamics of neoantigen presentation by the tumor cells demands high peptide purity and yield, and also the ability to quickly synthesize many peptides in parallel for timely treatment <GPT Neoantigen whitepaper, March 2020>. Such performance will be invaluable in fighting COVID-19.
In the highly dynamic world of COVID-19, a snapshot reveals the value of peptides in fighting the disease.
SARS-CoV-2 infection, like SARS-CoV that caused the SARS outbreak in 2003, invades host cells by binding the human angiotensin-converting enzyme 2 (ACE2) receptor on the cell surface through its viral spike protein (S; Figure 1), and it may be possible to develop peptide therapeutics that disrupt this protein-protein interaction (PPI).
For example, pre-incubation with two peptides reduced the infectivity of SARS-CoV in cell culture by over 10,000-fold, and infectivity was completely inhibited by combining three peptides (4). A similar method involves a 23-aa peptide fragment of the ACE2 peptidase domain α1 helix that binds to the S receptor binding domain of SARS-CoV-2 with low nanomolar affinity that could prevent the entry of virus into human cells (5, 6).
In another approach, bioinformatics has been used to search for targets of immune responses to SARS-CoV-2 (7). Multiple specific regions in SARS-CoV-2 have been identified that have high homology to the SARS-CoV virus and could be promising targets to guide peptide vaccine design.
The search and discovery of potential therapeutic peptides and epitopes for vaccine design puts a number of demands on the peptide sequence:
To illustrate how peptide synthesis can meet these needs, 24 peptides from a study on epitopes for COVID-19 vaccine development (7) were synthesized in parallel, followed by optimization of the synthesis of specific sequences.
The synthesis by Gyros Protein Technologies of the 24 SARS-CoV-2 peptides (Table 1) with therapeutic potential published by Grifoni and colleagues (7) represents a real-world example of synthetic challenges brought by a diverse range of sequences:
The steps in this process included:
PV1
EVPVAIHADQLTPTWRVYSTGS
RV1
DAVDCALDPLSETKCTLKSFTVEKGIYQTSN
PV2
FSQILPDPSKPSKRSFIE
RV2
FGAGAALQIPFAMQMAYRFNGI
PV3
PLLESELVIGAVILRGHLRI
RV3
MADSNGTITVEELKKLLEQWNLVI
PV4
NNNAATVLQLPQGTTLPKGF
RV4
RPQGLPNNTASWFTALTQHGK
PV5
IRGWIFGTTLDSKTQSLL
RV5
NKHIDAYKTFPPTEPKKDKKKKTDEAQPLPQRQKKQPTVTLLPAADM
PV6
QPFLMDLEGKQGN
RV6
CTFEYVSQPFLMD
PV7
KSFTVEKGIYQTSNFRVQ
RV7
TRFQTLLALHRSYLTPGDSSSGW
PV8
KLPDDFTGCV
RV8
SASFSTFKCYGVSPTKL
PV9
YLYRLFRKSNLKPFERDI
RV9
NLDSKVGGNYNYLYRLFR
PV10
KPFERDISTEIYQ
RV10
QSIIAYTMSLGAENSVAY
PV11
SIIAYTMSL
RV11
TECSNLLLQYGSFCTQL
PV12
DSLSSTASALGKLQDVV
RV12
VKQIYKTPPIKDFGGFNF
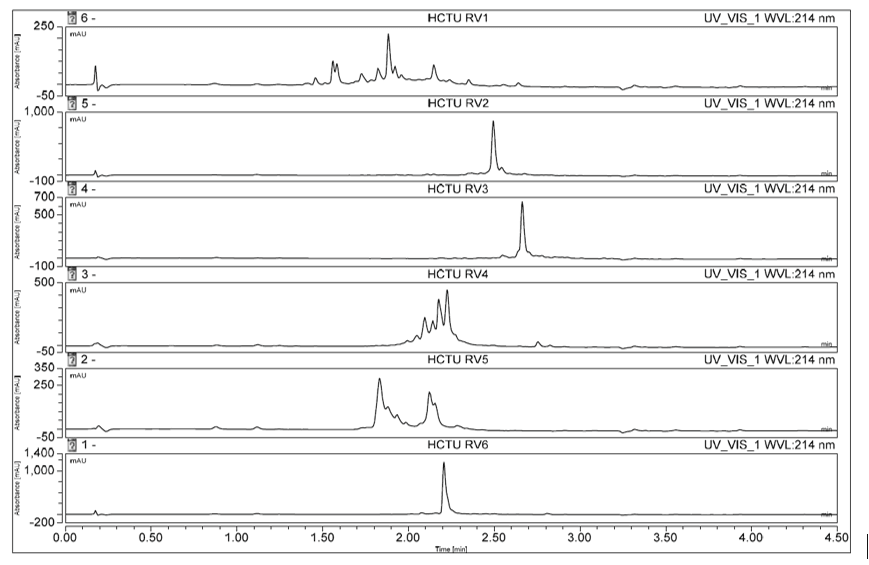
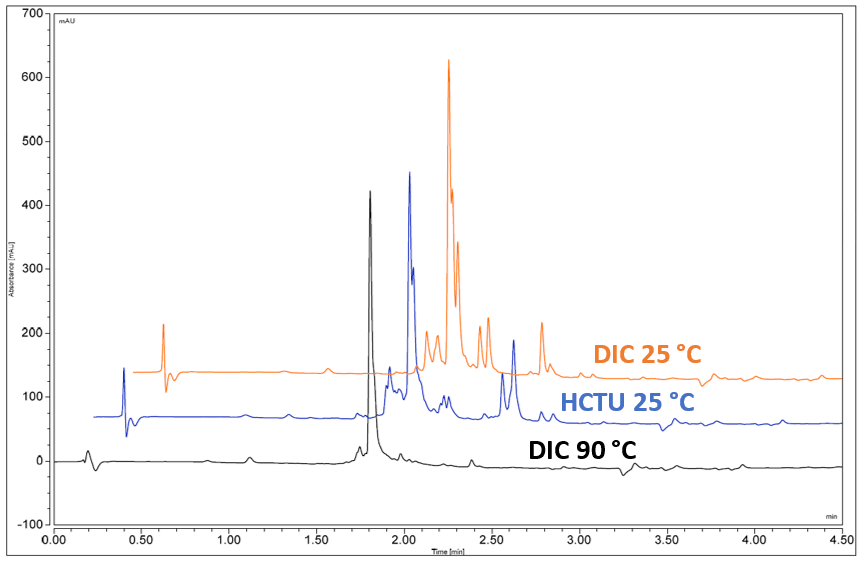
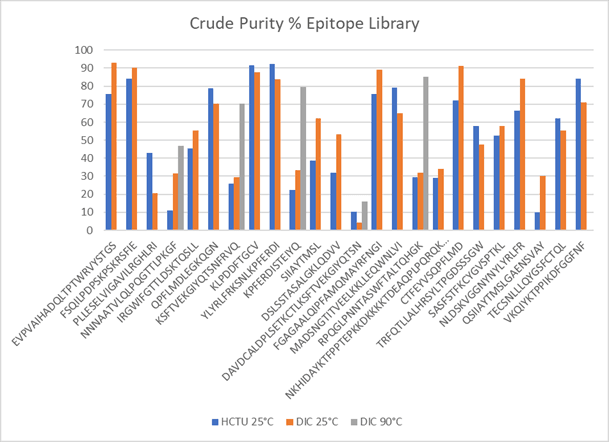
The first step in parallel synthesis on Symphony® X used fast HCTU/DIPEA and capping chemistry at 25 °C that enabled short coupling times (5 min), with the longest peptide (48-mer) taking 33 hours to synthesize. This chemistry resulted in crude purities of, for example, six of the 24 peptides in the range 17.9–79.1% (Fig. 2; full data for all peptides is shown in Fig. 5).
Figure 2. Analysis by reverse phase HPLC revealed that the crude purity of six of the 24 peptides, RV1–RV6, synthesized using HCTU/DIPEA chemistry ranged from 17.9% up to 79.1%.
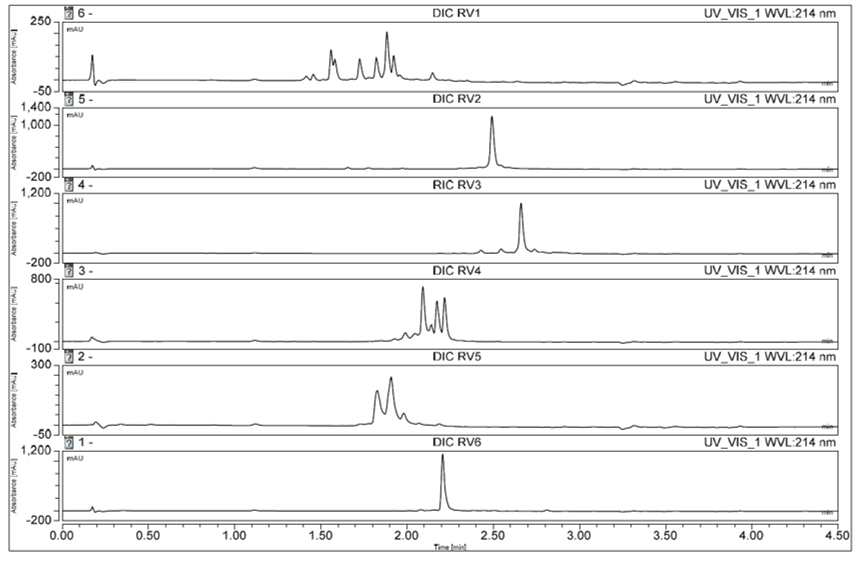
The second round involved exploiting the capacity of Symphony X to screen different chemistries to find optimum conditions, and involved testing the efficiency of DIC/OxymaPure coupling chemistry for all 24 peptides at 25 °C.
The results were slightly better compared to the syntheses based on HCTU in the first round, but the reaction time for DIC/OxymaPure is longer, which resulted in a longer overall cycle time. Some peptides did not perform well with either chemistry at 25 °C, therefore the next step was to try raising the coupling temperature.
Figure 3. Changing to DIC/OxymaPure chemistry increased the crude yield for some sequences.
In the third round of optimization, particularly challenging peptides were synthesized on PurePep® Chorus using DIC/OxymaPure chemistry at 90°C.
Raising the temperature from 25 °C to 90 °C, using PurePep Chorus, improved crude purity for a number of peptides (Fig. 4):
PV4 NNNAATVLQLPQGTTLPKGF: 31.6 → 47.0%
PV7 KSFTVEKGIYQTSNFRVQ: 29.6 → 70.3%
PV10 KPFERDISTEIYQ: 33.3 → 79.5%
RV4 RPQGLPNNTASWFTALTQHGK: 31.8 → 85.1%
Figure 4: Optimizing the synthesis of PV7, for example, involved testing the chemistry from (Round 1) HCTU/DIPEA at 25 °C on Symphony X (25.8 % crude purity) and (Round 2) DIC/OxymaPure (29.6%), and then (Round 3) increasing the reaction temperature to 90 °C (70.3 %), run on PurePep Chorus.
Figure 5. Summary of optimizing the synthesis of a diverse range of 24 peptides in an epitope library. The initial coupling reagent scan compared HCTU/DIPEA with DIC/OxymaPure chemistry at 25 °C using Symphony X. The crude purity of some problematic peptides was improved by synthesis at 90 °C with DIC/OxymaPure chemistry using PurePep® Chorus.
This Case Study represents a typical real-world example of the challenges that must be overcome when synthesizing a library of peptides with highly diverse sequences (Fig. 5).
Abbreviations
DIC - N,N' -Diisopropylcarbodiimide
DIPEA - N, N-Diisopropylethylamine
HCTU - 2-(6-Chloro-1H-benzotriazole- 1-yl)-1,1,3,3-tetramethyluronium hexafluorophosphate.
Researching virus infection and developing therapeutics and vaccines requires a lot from a peptide synthesizer. Symphony® X peptide synthesizer with the proprietary PurePep® Pathway, has the ability to run 12 independent reaction vessels and 24 reaction vessels overall, which is often desirable when dealing with peptide libraries.
Speed – able to synthesize 24 ‘epitope-type’ peptides in 12 hours (15–20 mers).
Purity is ensured due to the focus on ‘right first time’ methodology, with no reagent or resin cross-contamination, and minimizes the risk of re-synthesis.
Unique in providing multi-channel synthesis under inert atmosphere and with no cross contamination.
Supports GMP manufacture, designed for 21 CFR part 11 compliance, together with IQ/OQ support and PQ guidance are available to support work in regulated environments.
The proprietary PurePep® Pathway comprises fluidics that minimize cross-contamination, dead volumes, and reagent carryover. This is especially crucial for the synthesis of long sequences, in which even small amounts of impurities, side products, and incomplete reactions over many cycles can drastically reduce the final purity and yield of desired peptides.
Aggregation, secondary structure, steric hindrance, and conformational effects can still pose challenges in synthesis, and PurePep® Chorus features enabling technologies that aid the synthesis of complex peptides and peptidomimetic sequences. Intellisynth™ real-time UV monitoring optimizes reaction times to ensure complete deprotection. By monitoring at 301 nm, the instrument measures the progress of the reaction – avoiding guesswork that can lead to incomplete deprotections, deletions, and side reactions. This feature is available on all reaction vessels, providing UV monitoring on up to six peptides in a single synthesis.
A modular peptide synthesizer that is in-lab upgradeable, from 2 to 4 to 6 reaction vessels to meet productivity needs.
Independent induction heating, simultaneous and configurable to multiple vessels, and the ability to run multiple conditions in one run speeds up method development.
Icon-driven intuitive software platform with pre- programmed methods, ability to import sequences and reagent preparation calculators.
Designed for 21 CFR part 11 compliance, together with IQ/OQ support and PQ guidance are available to support work in regulated environments.
References
1. The COVID-19 vaccine development landscape. Thanh Le T et al, Nat Rev Drug Discov. 2020 Apr 9. doi: 10.1038/d41573-020-00073-5. 2. Screening 10,000 compounds identifies six potential COVID-19 therapeutics. https://www.drugtargetreview.com/news/60316/screening-10000-compounds-identifies-six-potential-covid-19-therapeutics/ 3. Biopharma products in development for COVID-19. https://www.bioworld.com/COVID19products 4. Synthetic peptides outside the spike protein heptad repeat regions as potent inhibitors of SARS-associated coronavirus. Zheng B-J et al. Antiviral Therapy 10 393-403. 2005 5. The first-in-class peptide binder to the SARS-CoV-2 spike protein. Zhang G. et al. bioRxiv preprint doi: https://doi.org/10.1101/2020.03.19.999318.
6. An experimental peptide could block Covid-19. http://news.mit.edu/2020/peptide-drug-block-covid-19-cells-0327 7. A Sequence Homology and Bioinformatic Approach Can Predict Candidate Targets for Immune Responses to SARS-CoV-2. Grifoni, A. et al. Cell Host & Microbe 27, 671–680 April 8, 2020 https://doi.org/10.1016/j.chom.2020.03.002
Website: www.gyrosproteintechnologies.com
Twitter: @GyrosProteintec
LinkedIn: Gyros Protein Technologies AB