Nejc Pavlin, Blaž Bakalar, Janja Skok, Špela Kralj, Andreja Gramc Livk, and Aleš Štrancar BIA Separations, now a Sartorius company
Plasmid DNA (pDNA) has become a crucial component in the production of next generation therapeutics such as messenger RNA (mRNA) and viral vectors.
As companies ramp up their production capabilities and move towards clinical applications, obtaining cGMP grade pDNA has become a production bottleneck, leading to lengthy production delays.
There is a growing market demand for solutions that can streamline the production of cGMP pDNA and help optimize down-stream processes (DSP) for better yields & purity.
The key step in this process is having quantifiably reliable analytics that give rapid results for process optimization and scale-up, as well as production runs.
Establishing and expanding in-house pDNA production platform in a quick and efficient manner will be a key differentiator between more and less successful next generation therapeutics projects.
Traditionally plasmids have been used for transfection and viral vector production [1] [2], as well as a direct therapeutic agent [3].
Recently, however, with the emergence of mRNA as the modality of choice for many new and developing therapies the demand for cGMP grade pDNA has grown dramatically.
This is mainly due to the key role of linearized plasmids as in vitro transcription (IVT) reaction templates [4].
While some applications of pDNA have similar general purity requirements, e.g. hcDNA/RNA/protein/endotoxin content, others have specific purity requirements, e.g. super coiled (SC)/open circular (OC)/linear (LIN)/multimer isoform pDNA separation & quantification [5].
In all cases, the host cell related impurities should be monitored and their removal optimized (hcDNA/RNA/protein/endotoxin).
Production of therapeutic plasmids and viral vectors requires monitoring and removal of process related impurities.
PATfix™ is biocompatible HPLC analytical system, specifically designed for rapid analysis and in-process control of gene therapy products.
The PATfix pDNA platform, optimized for pDNA analytics, allows users to perform analytics in a rapid at-line fashion, while minimizing the requirement for skilled operators.
This is made possible by built-in analytical methods, step-by-step user guides, and providing a certified pDNA standard, streamlining identification and quantification of bioprocess species.
Herein, we present lysis optimization of pFix5 plasmid obtained from raw E. coli paste, and subsequently its linearization suitable for IVT reaction template.
Specifically, we show the advantages to be gained during process development and production by using the PATfix pDNA platform to rapidly separate and quantify all key components, thus optimizing the plasmid processing workflow.
For successful IVT reaction it is critical to remove all the residual RNA, which would adversely affect the IVT reaction, generating off-target impurities that would be hard to separate from the targeted mRNA.
All experiments were performed using pFix5 crude biomass from E. Coli and pFix5 plasmid standard (4.7 kb, BIA Separations).
Analytical chromatographic experiments were performed on a PATfix HPLC system (Figure 1) composed of two analytical pumps, a multiwavelength UV-Vis detector equipped with a 10 mm flow cell. PATfix software was used for data analysis.
Chromatography separation was achieved using a CIMac™ pDNA (DEAE weak anion exchanger, 0.3 mL, 1.4 µm channels) monolith column from BIA Separations [6].
Loading buffer was composed of 100 mM TRIS, 300 mM guanidine chloride, 1 % Tween-20 at pH 8.0, and elution buffer was composed of 100 mM TRIS, 300 mM guanidine chloride, 700 mM NaCl, 1 % Tween-20 at pH 8.0 [7].
Other experimental details are included in the discussion below. HPLC analysis was orthogonally checked using agarose gel electrophoresis (AGE).
Figure 1: PATfix HPLC platform
Different methods of pDNA extraction have been described in the literature; among them is alkaline lysis, which is currently the most commonly used [8].
Although alkaline lysis is an established method for pDNA isolation, some drawbacks are recognized.
They include the capture of pDNA in cellular residues, leading to lower pDNA yields, lengthy procedure, and an increase in volume due to the buffers used leading to an increase in production costs [9].
Optimizing alkaline lysis parameters is a key to increase pDNA yield, shorten the time required for alkaline lysis, and reduce production costs [10].
In this work, two crucial lysis parameters were optimised. These include the degree of resuspension of the cell biomass before alkaline lysis, and the amount of added CaCl2 after neutralization of the alkaline lysate.
The resuspension buffer contained 50 mM TRIS, 10 mM EDTA pH 8.0. The effect of resuspension on the amount of isolated pDNA and RNA was studied at three different stages of resuspension; 10x, 30x and 50x.
For alkaline lysis, an equal volume of lysis buffer (0.2 M NaOH, 1 % SDS) was added to the resuspended biomass sample and incubated for 5 min.
After lysis, the sample was neutralized by the addition of 3 M CH3COOK (volume equal to the volume of lysis buffer).
Different amounts of 5 M CaCl2 were then added to the neutralized sample so that the final concentration of CaCl2 in the sample was 0, 0.25 M CaCl2, 0.5 M CaCl2, 0.75 M CaCl2 and 1 M CaCl2.
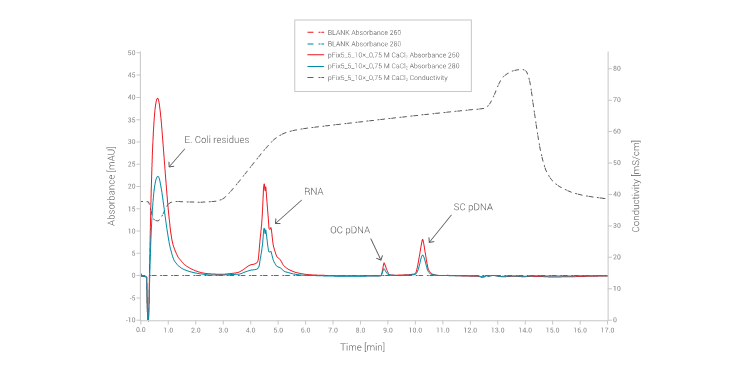
All samples were centrifuged and diluted 100x before HPLC analysis. In Figure 2 typical chromatogram obtained with HPLC analysis is shown. Using NaCl as the eluent, proteins and other E. coli residues do not bind to the CIMac pDNA column.
In contrast, RNA and pDNA moieties are efficiently captured on the weak anion exchanger monolith and are eluted in two linear gradients. Additionally, CIMac pDNA columns can also separate different pDNA isoforms and facilitate accurate determination of their ratio.
Figure 2: Typical chromatogram of centrifuged neutralised lysate recorded using PATfix pDNA analytics
The PATfix pDNA platform has proven to be a powerful tool for monitoring and optimizing the alkaline lysis of E. coli cells.
The CIMac pDNA column efficiently separates RNA and pDNA isoforms from undesirable moieties even in crude cell lysates.
This enables qualitative and quantitative determination of pDNA isoforms and impurities during the whole DSP, all in just a 17 min run.
Figure 3: Lysis optimization results using PATfix HPLC CIMac pDNA analytics
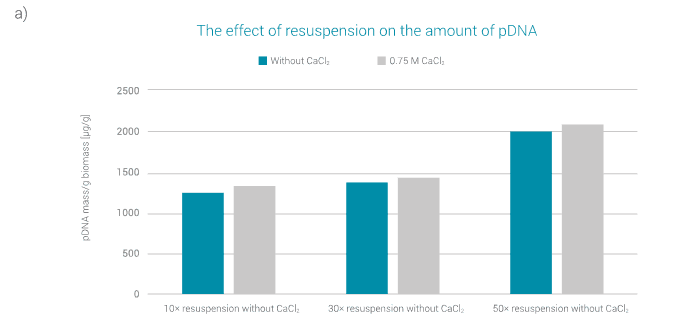
Figure 3a shows absolute amounts of pDNA as determined by PATfix pDNA analysis, depending on the factor of resuspension (10x, 30x, 50x) and the addition of CaCl2 (0.75 M).
The results show that the factor of resuspension buffer has a strong influence on the amount of extracted plasmid. The amount of extracted plasmid increases with the increasing factor of resuspension buffer.
In addition, the amount of extracted plasmid is affected by the addition of CaCl2, but this effect is not as pronounced as in the case of the factor of resuspension buffer.
A slightly larger amount of plasmid was extracted by adding CaCl2.
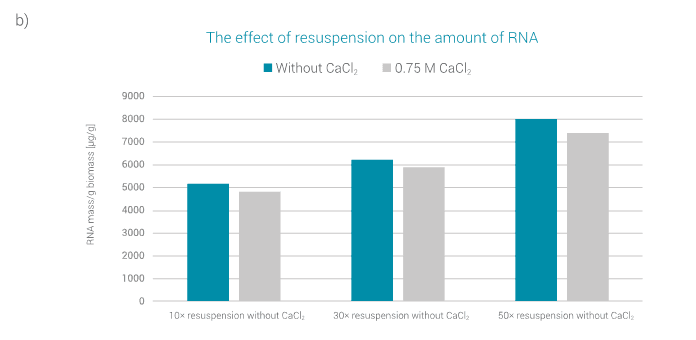
A similar trend can also be observed in the case of the amount of RNA extracted, as shown in Figure 3b. The amount of RNA extracted increases with a higher factor of resuspension.
The effect of CaCl2 addition differs with the amount of RNA. It is known from the literature [11] that CaCl2 causes precipitation of RNA, leading to a smaller amount of RNA in the analysed sample.
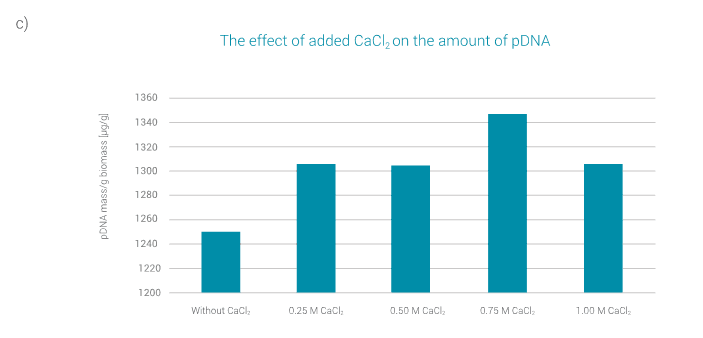
The amount of pDNA extracted from cell biomass as a function of the amount of added CaCl2 is shown in Figure 3c. The resuspension rate was the same in all cases; 10x.
The lowest amount of extracted plasmid is obtained when CaCl2 is not added to the sample.
The best plasmid extraction yield is obtained in the case of the addition of 0.75 M CaCl2. Larger amounts of added CaCl2 negatively affect the amount of plasmid extracted.
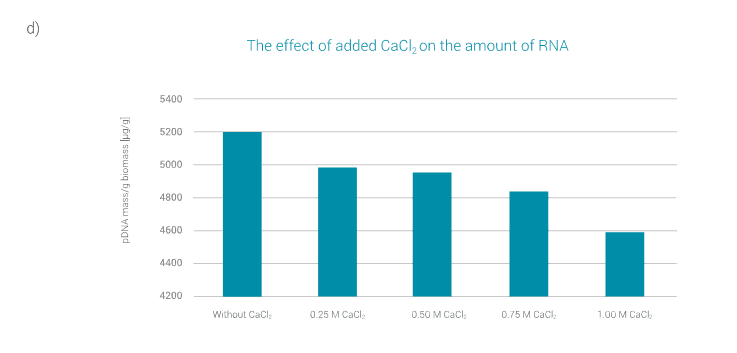
As the amount of CaCl2 changes, so does the amount of extracted RNA. As can be seen in Figure 3d, the amount of extracted RNA decreases with the amount of CaCl2 added.
The lowest content of extracted RNA is obtained when 1 M CaCl2 is added to the neutralised lysate sample after neutralization.
Based on the optimization results, it can be concluded that the most suitable conditions for alkaline lysis, in terms of resuspension and addition of CaCl2, are at 50x resuspension and the addition of 0.75 M CaCl2.
The addition of 1 M CaCl2 results in less RNA in the neutralised lysate sample, but the amount of extracted plasmid is also reduced at this CaCl2 concentration.
Beside alkaline lysis and pDNA purification, plasmid linearization is the first key step in mRNA production that benefits from PATfix pDNA analytics and associated process optimization.
Due to the high cost of restriction enzymes and the plasmid itself, it is crucial that the linearization process is optimized in terms of increasing the quality of the final product while reducing production costs.
Here, we present a case study for monitoring plasmid linearization kinetics using PATfix pDNA analytics.
Figure 4: a) Monitoring pFix5 plasmid linearization using PATfix pDNA analytics, b) Relative proportion of pDNA isoforms vs. time
For linearization of pFix5 plasmid, NdeI (NEB) enzyme was used.
Linearization mixture was composed of linearization buffer (500 mM KCOOCH3, 200 mM Tris, 100 mM Mg(COOCH3)2, pH 7.9), pFix5 plasmid, NdeI enzyme, using UltraPure™ Dnase & Rnase – Free Distilled Water.
Linearization mixture was incubated at 37°C, and at different time intervals, with aliquots quantitatively sampled.
Aliquots were immediately inactivated using EDTA (final concentration in aliquot) and analysed using PATfix pDNA analytics.
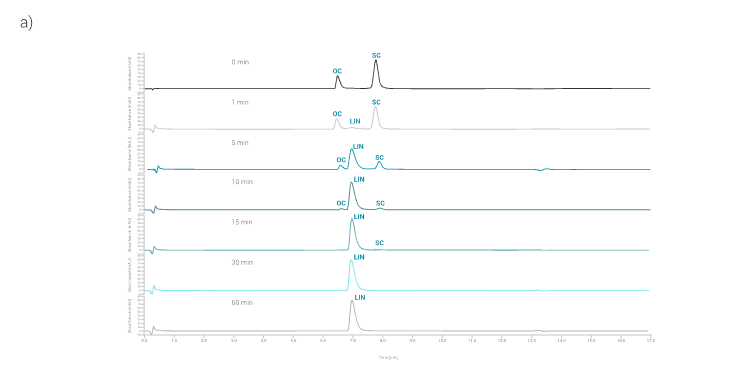
Figure 4a shows pFix5 plasmid linearization at different reaction times. Linearization starts to take place in the first minute of the linearization reaction.
After 15 min of linearization, almost all of the pDNA is converted to a linearized isoform (decrease of peaks for OC and SC and increase of peaks for LIN pDNA).
After 30 min, the linearization reaction is completed, as the peaks belonging to the OC and SC isoforms of pDNA are no longer detected by HPLC analysis.
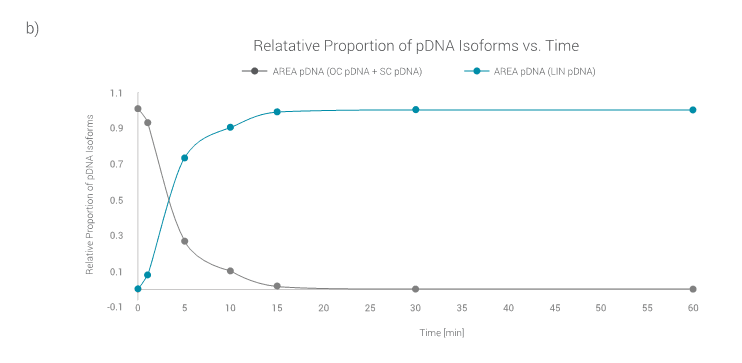
Only the peak characteristic for the LIN pDNA isoform is detected. This is nicely displayed in Figure 4b, where the relative proportion of pDNA isoforms are shown.
After 15 min of linearization reaction, the relative proportion of OC and SC pDNA isoform, as well as LIN pDNA isoform, reach a plateau, at 0 and 1 respectively.
This indicates that linearization reaction comes to an end and that both OC and SC isoforms were completely converted to the LIN pDNA isoform.
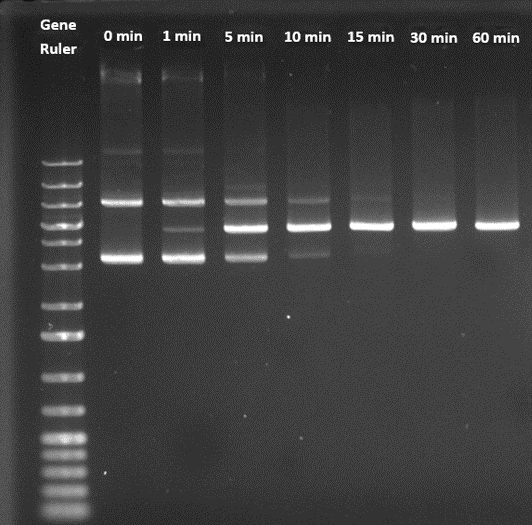
Figure 5: Monitoring linearization process using Agarose gel electrophoresis (AGE)
The time-course pDNA linearization results obtained from the PATfix pDNA analysis were orthogonally checked by AGE (Figure 5).
From Figure 5 is observed that the linearization reaction starts as soon as the enzyme is added to the linearization reaction mixture.
The linearization process is completed in 30 min, since no more bands representing the OC and SC isoform of pDNA are observed.
While the reaction in this case is fast, another potential avenue of optimization, especially for larger scales, is the enzyme usage, allowing the user to optimize for lower the enzyme usage while lengthening the reaction.
Sensitivity of PATfix pDNA analytics is especially useful for such process optimization, where very low limit of detection (LOD) and limit of quantification (LOQ) capability is required in order to obtain meaningful results.
The results obtained by AGE analysis were in line with the PATfix pDNA analytics.
The advantage of PATfix pDNA analytics is mainly in terms of sensitivity and improved detection of RNA impurities, while the quantification is an additional benefit, something that AGE struggles with.
PATfix pDNA analytics allow efficient qualitative and quantitative monitoring of pDNA linearization.
It enables estimation of the kinetics of plasmid linearization, which allows us to optimize the linearization process to reduce required time and cost.
[1] D. E. Andreev, I. M. Terenin, S. E. Dmitriev, and I. N. Shatsky, “Pros and cons of pDNA and mRNA transfection to study mRNA translation in mammalian cells,” Gene, vol. 578, no. 1, pp. 1–6, Mar. 2016, doi: 10.1016/j.gene.2015.12.008.
[2] R. J. Anderson and J. Schneider, “Plasmid DNA and viral vector-based vaccines for the treatment of cancer,” Vaccine, vol. 25, no. SUPPL. 2, pp. 24–34, 2007, doi: 10.1016/j.vaccine.2007.05.030.
[3] M. A. Liu, “DNA vaccines: a review,” J. Intern. Med., vol. 253, no. 4, pp. 402–410, Apr. 2003, doi: 10.1046/j.1365-2796.2003.01140.x.
[4] N. Pardi, M. J. Hogan, and D. Weissman, “Recent advances in mRNA vaccine technology,” Curr. Opin. Immunol., vol. 65, pp. 14–20, Aug. 2020, doi: 10.1016/j.coi.2020.01.008.
[5] F. Smrekar et al., “Preparation of pharmaceutical-grade plasmid DNA using methacrylate monolithic columns,” Vaccine, vol. 28, no. 8, pp. 2039–2045, 2010, doi: 10.1016/j.vaccine.2009.10.061.
[6] B. Gabor, U. Černigoj, M. Barut, and A. Štrancar, “Reversible entrapment of plasmid deoxyribonucleic acid on different chromatographic supports,” J. Chromatogr. A, vol. 1311, pp. 106–114, Oct. 2013, doi: 10.1016/j.chroma.2013.08.075.
[7] U. Černigoj et al., “Guanidine Improves DEAE Anion Exchange-Based Analytical Separation of Plasmid DNA,” Electrophoresis, pp. 1–22, 2021, doi: 10.1002/elps.201400126.
[8] H. C. Birnboim and J. Doly, “A rapid alkaline extraction procedure for screening recombinant plasmid DNA,” Nucleic Acids Res., vol. 7, no. 6, pp. 1513–1524, 1979.
[9] S. Haberl, M. Jarc, A. Štrancar, M. Peterka, D. Hodžić, and D. Miklavčič, “Comparison of Alkaline Lysis with Electroextraction and Optimization of Electric Pulses to Extract Plasmid DNA from Escherichia coli,” J. Membr. Biol., vol. 246, no. 11, pp. 861–867, Nov. 2013, doi: 10.1007/s00232-013-9580-5.
[10] A. Abdulrahman and A. Ghanem, “Recent advances in chromatographic purification of plasmid DNA for gene therapy and DNA vaccines: A review,” Anal. Chim. Acta, vol. 1025, pp. 41–57, Sep. 2018, doi: 10.1016/j.aca.2018.04.001.
[11] A. Eon-Duval, K. Gumbs, and C. Ellett, “Precipitation of RNA impurities with high salt in a plasmid DNA purification process: Use of experimental design to determine reaction conditions,” Biotechnol. Bioeng., vol. 83, no. 5, pp. 544–553, Sep. 2003, doi: 10.1002/bit.10704.
Nejc Pavlin is the project manager, Blaž Bakalar is the PATfix product manager, Špela Kralj is the master student, Janja Skok is the associate scientist,
Andreja Gramc Livk is the head of process analytics development, and Aleš Štrancar is the CEO of BIA Separations, now a Sartorius company.
The authors gratefully acknowledge valuable contributions of Urh Černigoj, Rok Sekirnik and Kaja Tudja of BIA Separations.
Visit us at our website
Download our brochure